
Evolución genómica. Últimos 20 años
En estos últimos 25 años los laboratorios de genética o biología molecular se han transformado radicalmente y muchas poyatas de laboratorio se han convertido en mesas con ordenadores conectados a internet. Las comparaciones de miles de secuencias para buscar mutaciones en un gen concreto, la búsqueda de posibles elementos reguladores comparando centenares de genomas secuenciados o la predicción de la estructura de proteínas que nunca han sido purificadas se han convertido en rutina. Y obtenemos respuestas casi instantáneas a preguntas que serían impensables hace pocos años. Hace menos de 25 años, en el año 2000, se publicó el primer genoma completo de una planta, Arabidopsis thaliana, y un año después se publicó el primer borrador del genoma humano. Y supuso una revolución. A muchos les costará de creer, pero poco antes de la publicación del genoma humano había apuestas en muchos laboratorios sobre el número de genes que finalmente tendría ¿40.000, 50.000, o quizás 80 o 100.000 genes? Era de esperar que los humanos tuviéramos muchos más genes que una mosca o que una planta, porque además de ser organismos más complejos, tenemos un genoma más grande. Y sin embargo… La publicación del genoma humano nos deparó auténticas sorpresas. La primera, que el número de genes anotados no superaba los 30.000, un número que dos años más tarde se rebajó a unos 20.000; y la segunda, que estos genes sólo ocupan una pequeñísima parte del genoma. Por ello, El País, dando a conocer estos resultados, publicó un artículo en febrero de 2001 con el expresivo título de “El 95% del genoma humano es basura”. Esa basura genómica está formada, en su mayor parte, por transposones.
El papel de los transposones ha sido siempre controvertido desde que Barbara McClintock propusiera su existencia en los años 50 del siglo pasado, y han sido considerados durante mucho tiempo como basura genómica. Sin embargo, de forma paulatina se han ido acumulado evidencias de su importancia en la evolución de los genomas y en la regulación génica. Sólo 10 años después del artículo mencionado, en septiembre de 2012, el mismo periódico El País publicaba un artículo titulado “La basura genómica ya es medicina”, donde se explicaba que muchos de los elementos reguladores humanos se encuentran en secuencias de elementos móviles.
En este artículo proponemos un breve viaje en la historia para entender hasta qué punto las ideas de McClintock fueron revolucionarias en su tiempo y como fueron confirmadas con el avance de la genómica, para después hacer un resumen de lo que conocemos actualmente sobre el impacto de los transposones en los genomas eucariotas.
Los transposones, una idea visionaria de Barbara McClintock
Barbara McClintock (Figura 1) desarrolló con gran habilidad la citogenética de maíz a finales de los años 40 del siglo pasado. McClintock recopiló datos de numerosos cruces genéticos y observó detalladamente los diferentes caracteres de los granos de las mazorcas y sus coloraciones en las plantas hijas. En estos estudios encontró las primeras pruebas físicas de la recombinación génica, cuya existencia habían sugerido Sturtevant y Morgan en los años veinte, además de las primeras pruebas de la transposición. McClintock descubrió que en los cromosomas de maíz había determinantes genéticos (el concepto de gen era aún muy debatido) que producían roturas en los cromosomas, de aquí el nombre Disociador, (Dissociator, Ds por sus siglas en inglés) y vio que éstos podían cambiar posiciones en el genoma. También encontró que había otros determinantes genéticos que parecían influenciar (controlar) el comportamiento de Ds, a los que llamó Activadores (Activators, Ac por sus siglas en inglés). Estos estudios dieron lugar a dos ideas importantes: (1) los determinantes genéticos podían moverse de sitio en los cromosomas (transposición); (2) estos determinantes controlaban la expresión génica.

Barbara McClintock (1902-1992), Department of Genetics, Carnegie Institution at Cold Spring Harbor, New York, trabajando en su laboratorio (fuente Smithsonian Institution, http://commons.wikimedia.org/).
En 1951, en el congreso anual de Cold Spring Harbour, Barbara McClintock presentó por primera vez las evidencias experimentales de la existencia de los transposones. En su exposición McClintock relacionó también este descubrimiento con la idea del control génico de la expresión al bautizar los elementos móviles como elementos controladores “controlling elements” del desarrollo de la planta. Es difícil hacerse una idea del salto conceptual que significaba el descubrimiento de la transposición hoy en día. Que el ADN era la molécula encargada de transmitir la herencia se demostró un año después (1952, Hersey y Chase) y la estructura de la molécula del ADN dos años más tarde (1953, Watson y Crick). Tuvieron que pasar aún diez años, hasta 1961, para que se asociara por primera vez un triplete de bases a un codón, y cinco más, hasta 1966, para descifrar todo el código genético. Todos estos descubrimientos fueron muy rápidos y pedían tiempo para su asimilación. Incluso el concepto de gen estaba en debate. Probablemente añadir la idea de movilidad dentro del ADN creaba demasiada confusión para que la transposición pudiera ser rápidamente aceptada.
Durante las siguientes décadas, se descubrieron nuevos elementos transponibles en maíz (en 1955 ya se habían identificado, Spm, Dotted, Modulator y Enhancer), en Drosophila (1965), en fagos (1972), bacterias (1966) y finalmente en levadura (1979) demostrando que los transposones eran parte intrínseca de los genomas procariotas y eucariotas, y mostrando la gran variedad estructural de estos elementos (Figura 2) que se corresponde con distintos mecanismos de transposición (Figura 3). En 1983, Barbara McClintock recibió el Premio Nobel de Fisiología o Medicina por el descubrimiento de los elementos móviles.

Clasificación general de los Transposones: clases y superfamílias principales. Las cajas azules representan las regiones codificantes para las distintas actividades necesarias para la transposición. Las flechas sólidas indican las repeticiones terminales directas (Long Terminal Repeat, LTR por sus siglas en inglés), o invertidas (Terminal Inverted Repeat, TIR por sus siglas en inglés). Las flechas finas indican las unidades transcripcionales.
Aunque fue muy polémica su definición de los elementos móviles como elementos controladores, cuando Jacob y Monod presentaron en 1960 su modelo del operón en E. coli, McClintock saltó de alegría al ver los paralelismos con su idea de “elementos controladores”. La idea de que existían los operones en bacterias y los elementos controladores en maíz, demostraba, según McClintock, que este sistema de regulación era extensible a todo organismo vivo. Es más, para McClintock, el control sobre la expresión génica debía de estar conectado con una causa celular (algo en el citoplasma), o en el ambiente en el que se desarrolla el organismo. Poder responder frente a cambios en el ambiente (estrés) regulando la expresión génica permite a los organismos hacer frente a estos cambios, adaptarse, evolucionar. Como veremos en el apartado siguiente, la intuición de McClintock de la relación de los tranposones con la adaptación al estrés ha resultado certera.
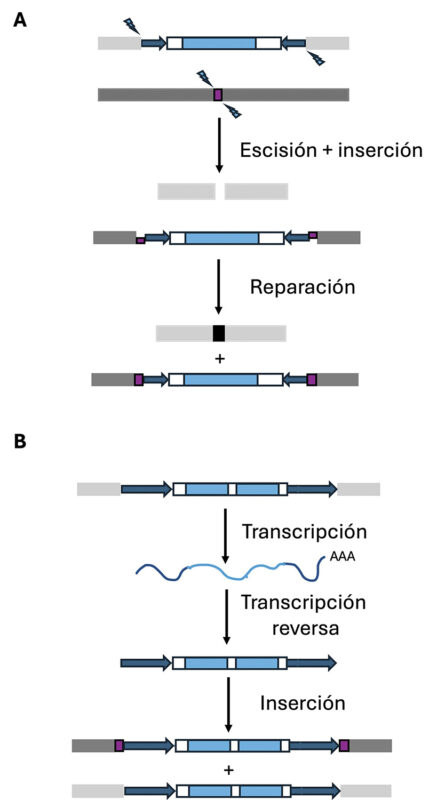
Principales mecanismos de transposición. Los transposones se representan siguiendo el mismo código que en la Figura 2. (A) Transposición de los elementos móviles de clase II, o transposones de ADN. Mecanismo conocido como “cortar y pegar”. Los sitios de corte de la transposasa en el locus dador (cajas gris claro) y aceptor (cajas gris oscuro) se muestran con flechas azules. El sitio de inserción (target site) que será duplicado a ambos lados de la inserción se muestra como una caja violeta. (B) Transposición de los elementos móviles de clase I, o retrotransposones. Mecanismo conocido como “copiar y pegar”. El sitio de inserción, que se repara como en el caso de los transposones de clase II y se verá duplicado como resultado de la inserción, se muestra como una caja violeta.
Riesgos y beneficios de un genoma con transposones: mutación, plasticidad y adaptación
Los transposones son elementos genéticos móviles que se encuentran repetidos en el genoma. Estas dos características, su movilidad y su repetitividad, los hacen altamente mutágenos. Y por ello, la presencia de un gran número de estos elementos en los genomas parece paradójica. ¿Cómo es posible mantener la información genética de célula a célula y de generación en generación, cuando esta información también contiene elementos que tienden a modificarla? En realidad, la información genética no se mantiene inalterable durante la vida de un organismo o la evolución de una especie, y tanto las mutaciones somáticas como las germinales son, de hecho, esenciales. La cuestión es cuánto cambio puede ser útil y cuándo empieza a ser un riesgo demasiado alto. Y esto puede depender del tipo de organismo, de la célula o el tejido en cuestión o de las condiciones en las que se encuentre el organismo. La actividad de los transposones no es igual en todas las células y, por ejemplo, suele estar fuertemente reprimida en la línea germinal, mientras que la actividad en ciertas células somáticas puede ser particularmente alta. Un ejemplo paradigmático es la activación de los retrotransposones L1 durante el desarrollo neuronal en humanos, que puede ser en parte responsable de la alta mosaicidad y plasticidad de las neuronas, características esenciales para el correcto desarrollo del cerebro.
Por otra parte, el trabajo de muchos laboratorios durante los últimos 20 años ha puesto de manifiesto que la actividad de los transposones de plantas está frecuentemente ligada a situaciones de estrés, confirmando una vieja hipótesis de McClintock que proponía a los transposones como elementos reguladores que permiten la reorganización del genoma en situaciones a las que el organismo no está bien adaptado. Muchos transposones se activan en situaciones de estrés, en particular en plantas, debido a que tienen promotores regulados por estrés. Por otra parte, trabajos recientes sugieren que algunos transposones podrían insertarse preferencialmente cerca de los genes de respuesta al estrés guiados por marcas epigenéticas específicas, lo que permitiría generar nueva variabilidad en estos genes y permitir al organismo explorar variaciones en la respuesta al estrés y adaptarse. De una manera general, la inserción de transposones, con sus promotores o elementos reguladores específicos, puede modificar la regulación de los genes que se encuentran cerca, y los datos acumulados en los últimos años sugieren la importancia de este fenómeno en la evolución de la regulación de la transcripción en eucariotas. Por ejemplo, más de la mitad de las regiones de cromatina abierta específica de primates, que potencialmente contienen elementos regladores específicos, están en secuencias de transposones y muchos de los sitios de unión de factores de transcripción en estas especies también están en secuencias de transposones. También en plantas se ha podido demostrar que los transposones pueden amplificar y redistribuir en el genoma sitios de unión de factores de transcripción, dotando de plasticidad y capacidad de evolución a las redes transcripcionales.
Los transposones han sido pues una fuente de variación genética muy importante en la evolución de los eucariotas. Por ejemplo, muchas de las mutaciones clave para la domesticación y mejora de las plantas cultivadas, son debidas a la inserción de transposones. De la arquitectura de la planta del maíz moderno a la diversidad de color de las uvas o de las naranjas sanguinas, de la forma de algunos tomates a la piel suave de las nectarinas, todos son fenotipos provocados por la inserción de transposones.
Pero, por supuesto, solo una pequeña parte de las mutaciones supone una ventaja fenotípica y los eucariotas han desarrollado sofisticados mecanismos de control para regular la actividad de los transposones. El principal mecanismo de control de los transposones es el silenciamiento génico, y en particular su metilación y heterocromatinización. De hecho, es probable que los mecanismos epigenéticos de control de la expresión génica apareciesen en la evolución por la necesidad de controlar a los elementos genéticos invasores del genoma, como los virus y los transposones. Por lo tanto, es en buena medida la presencia de transposones, a pesar del riesgo que entrañan, o en algunos casos debido a ese riesgo y a la necesidad de controlarlo, lo que ha permitido alcanzar la complejidad de los genomas eucariotas.
Además de forzar el desarrollo de sistemas de control, los transposones también han sido una fuente importante de nuevos genes y funciones. Los transposones codifican para distintas proteínas, necesarias para asegurar su movilidad, que han sido domesticadas repetidas veces durante la evolución para realizar otras funciones celulares. En animales, quizás el caso más evidente son las Sincitinas que están implicadas en el desarrollo de la placenta de los mamíferos y que derivan de genes Env de retrotransposones (retrovirus endógenos), domesticados independientemente en distintos linajes de mamíferos. En plantas, varios factores de transcripción, como FHY3 y FAR, que regulan la transducción de señal de la luz, derivan de transposasas y varias proteínas que participan en la regulación epigenética de genes y transposones, como MAIL1, MAIN y ALP1 derivan de distintas proteínas de elementos móviles.
Los transposones como componentes estructurales de los cromosomas
Además de las funciones que hemos descrito en el apartado anterior, los transposones también son esenciales para asegurar la estructura y la homeostasis de los genomas eucariotas. Los telómeros de los genomas eucariotas se mantienen en la mayoría de los casos por la acción de la telomerasa, que copia por transcripción reversa un molde de ARN, un proceso filogenéticamente relacionado con la transposición de algunos retrotransposones. Una relación todavía más evidente si se tiene en cuenta que Drosophila, que no tiene telomerasa, asegura la longitud de sus telómeros por la transposición específica de dos retrotransposones, HeT-A y TART, al extremo de sus cromosomas. Por otra parte, los transposones pueden ayudar a mantener la integridad de los cromosomas insertándose en roturas de la doble cadena de ADN (Double-Strand Breaks, DSB por sus siglas en inglés) ayudando a reparar el ADN.
Un caso muy particular de rotura y reparación del ADN es la recombinación V(D)J que se da en los linfocitos de los vertebrados, y que permite generar una gran variedad de anticuerpos, inmunoglobulinas y receptores de células T. Esta recombinación está mediada por las recombinasas RAG1 y RAG2 que se unen, cortan y recombinan secuencias de ADN específicas. Tanto estos enzimas como las secuencias reconocidas derivan directamente de antiguos transposones.
Por otro lado, la relación de los transposones con el centrómero, esencial para la estructura y la dinámica de los cromosomas, es también muy estrecha. Además del hecho de que la proteína CNP-B, importante para la estabilidad centromérica en mamíferos, sea una antigua transposasa, los transposones son parte integral de los centrómeros de la mayoría de eucariotas y en Arabidopsis se ha podido demostrar que el silenciamiento de los retrotransposones centroméricos mediado por pequeños ARNs es esencial para el correcto funcionamiento del centrómero. La función centromérica está muy conservada en los eucariotas, pero paradójicamente su secuencia es extraordinariamente variable. Las secuencias centroméricas suelen ser altamente repetitivas y la región funcionalmente activa se define por la presencia de CENH3, la variante centromérica de la histona H3. Sin embargo, la secuencia del ADN varía entre cromosomas del mismo individuo o entre variedades o especies cercanas. En plantas, los centrómeros suelen estar formados por repeticiones en tándem de satélites centroméricos (CEN178 en Arabidopsis), e inserciones de retrotransposones, aunque en algunas especies los centrómeros pueden estar formados únicamente por secuencias de retrotransposones. La acumulación de retrotransposones en los centrómeros y zonas pericentroméricas puede ser debido a que esas regiones tienen una baja densidad en genes y por lo tanto, las inserciones de transposones tienen un efecto menos deletéreo, y la presión de selección para eliminarlos es menor. Por otra parte, la baja recombinación de esas regiones hace más difícil su eliminación. Pero además de estos mecanismos que podrían facilitar la acumulación de retrotransposones en el centrómero, se ha demostrado que existen retrotransposones centrofílicos que se insertan específicamente en estas regiones. Los centrómeros se caracterizan por una evolución rápida de su secuencia, dentro de la misma especie y entre especies distintas. Se ha propuesto que los centrómeros de plantas podrían alternar de forma dinámica entre estar formados por repeticiones centroméricas o retrotransposones. Los retrotransposones centrofílicos podrían colonizar un centrómero basado en repeticiones centroméricas, pero a su vez, existen algunos datos que sugieren que la eliminación por recombinación de los retrotransposones podría dar lugar a nuevas repeticiones simples en tándem que podrían convertirse en nuevas repeticiones centroméricas, lo que proporcionaría un mecanismo para explicar la rápida evolución de estas secuencias.
Conclusión
Casi setenta y cinco años después de que McClintock propusiera la existencia de los transposones, y gracias a los datos acumulados por un gran número de laboratorios, podemos afirmar que estos elementos, lejos de ser sólo “basura genómica”, son esenciales para explicar la estructura y la dinámica de los cromosomas, y asegurar al mismo tiempo, la complejidad y la plasticidad de los genomas eucariotas.
Para leer más
- Cui X, Cao X. Epigenetic regulation and functional exaptation of transposable elements in higher plants. Current Opinion in Plant Biology 21 (2014) 83–88. https://doi.org/10.1016/j.pbi.2014.07.001
- Joly-Lopez Z, Bureau TE. Exaptation of transposable element coding sequences. Current Opinion in Genetics & Development 49 (2018) 34–42. https://doi.org/10.1016/j.gde.2018.02.011
- Lisch D. How important are transposons for plant evolution? Nature Review Genetics 14 (2013) 49–61. https://doi.org/10.1038/nrg3374
- Naish M, Henderson IR. The structure, function, and evolution of plant centromeres. Genome Research 34 (2024) 161–178. doi: 10.1101/gr.278409.123
- Wells JN, Feschotte C. A field guide to eukaryotic transposable elements. Annual Review of Genetics 54 (2020) 539–561. https://doi.org/10.1146/annurev-genet-040620-022145
