
Introducción
En la última década, los condensados biomoleculares han cambiado nuestra visión de la organización celular. Frente al modelo clásico basado en orgánulos con membrana o en un citoplasma homogéneo, hoy sabemos que existen compartimentos dinámicos sin membrana que organizan las reacciones celulares. Estos condensados, formados mediante separación de fases, permiten concentrar proteínas y ARN en microambientes específicos con propiedades propias.
En plantas, este campo ha crecido rápidamente. Los condensados participan en procesos clave como la expresión génica, la señalización hormonal, el metabolismo y la respuesta al estrés, lo que resulta fundamental en organismos sésiles que deben adaptarse rápidamente a cambios ambientales. Algunos están conservados en todos los eucariotas, mientras que otros son específicos de plantas y responden a estímulos concretos.
A pesar de los avances, su estudio sigue siendo complejo. Comprender su formación y función requiere integrar conceptos de biofísica con biología celular, y su análisis experimental es especialmente desafiante en plantas debido a características como la pared celular o la autofluorescencia de los cloroplastos. En conjunto, los condensados representan un mecanismo rápido y reversible que permite a las plantas reorganizar su actividad celular frente a condiciones cambiantes.
Los gránulos de estrés como nodos integradores de la respuesta celular en plantas
Entre los condensados biomoleculares mejor caracterizados se encuentran los gránulos de estrés (SGs, por sus siglas en inglés stress granules), presentes en todos los eucariotas. Se trata de condensados biomoleculares que se forman en respuesta a condiciones adversas y están compuestos principalmente por ARN mensajeros (ARNm) detenidos en la traducción, subunidades ribosomales pequeñas, proteínas de unión a poli(A) (PAB, por sus siglas en inglés), factores eucarióticos de iniciación de la traducción (eIFs, por sus siglas en inglés eukaryotic initiation factors) y otras proteínas de unión a ARN. Debido a su composición, históricamente se les atribuyó un papel central en la regulación postranscripcional y en el control de la traducción durante el estrés. Sin embargo, estudios recientes han demostrado que también incorporan proteínas implicadas en la señalización, el metabolismo y otros procesos celulares, lo que ha llevado a redefinirlos como centros multifuncionales capaces de coordinar distintas respuestas celulares.
En el contexto de las plantas, esta plasticidad funcional adquiere especial relevancia. Factores como el aumento de temperaturas, la sequía o la salinidad, cada vez más intensos y frecuentes debido al cambio climático, exigen respuestas rápidas y eficientes. En este sentido, los SGs se perfilan como nodos dinámicos que integran señales de estrés y reorganizan la actividad celular, contribuyendo a la adaptación de las plantas a entornos cambiantes. Por ejemplo, una de sus funciones mejor establecidas es la regulación del destino del ARN. Durante el estrés, los SGs secuestran ARNm cuya traducción se ha detenido, protegiéndolos y permitiendo su reutilización cuando las condiciones mejoran (Figura 1). Este proceso no es aleatorio, ya que ciertos transcritos relacionados con la respuesta al estrés se estabilizan selectivamente en estos gránulos, favoreciendo una respuesta adaptativa más eficiente.
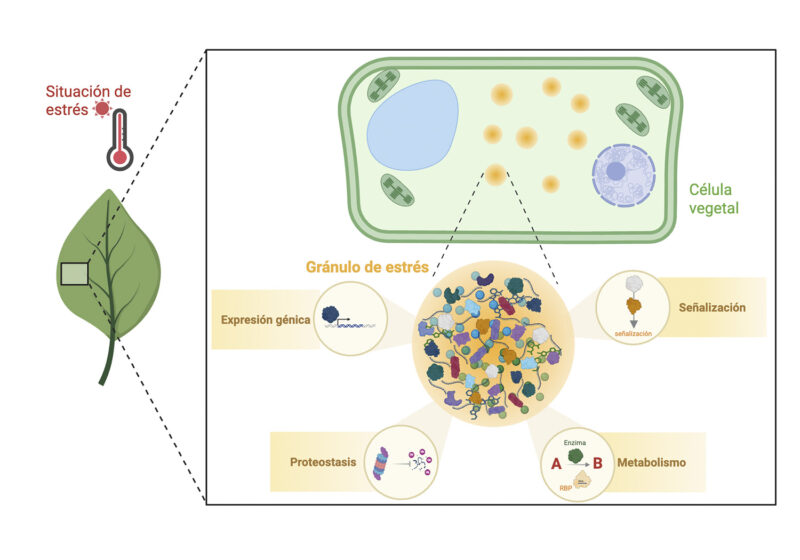
El gránulo de estrés como nodo integrador en plantas. Ante situaciones de estrés ambiental (p. ej. térmico), las células vegetales forman condensados dinámicos llamados gránulos de estrés (SGs, por sus siglas en inglés). Lejos de ser meros almacenes de ARN, estos gránulos actúan como centros multifuncionales que interconectan la expresión génica, la señalización, el metabolismo y la proteostasis celular, permitiendo una respuesta adaptativa rápida y eficiente.
Además, los SGs están estrechamente vinculados a otros procesos celulares (Figura 1). En el ámbito del metabolismo, se ha observado que enzimas implicadas en la detoxificación de especies reactivas de oxígeno (ROS, por sus siglas en inglés) o en rutas metabólicas centrales pueden reclutarse hacia estos condensados, modulando su actividad y contribuyendo al equilibrio redox durante el estrés. Esto sugiere que los SGs no sólo almacenan componentes, sino que reorganizan activamente la maquinaria metabólica. Otro aspecto relevante es su papel en la regulación de la homeostasis proteica. Durante condiciones adversas, estos condensados pueden secuestrar proteínas mal plegadas o componentes de los sistemas de degradación, lo que contribuye a prevenir la formación de agregados tóxicos y facilita la recuperación celular. Los SGs también actúan como nodos de señalización. Proteínas clave de vías de señalización, como quinasas implicadas en el balance energético celular, se localizan en estos condensados, donde su actividad puede ser modulada. De este modo, los SGs participan en la integración de señales que controlan la respuesta global al estrés (Figura 1).
En conjunto, estos hallazgos sustentan la visión de que los gránulos de estrés funcionan como centros organizadores que interconectan la regulación del ARN, el metabolismo, la señalización y la proteostasis. Esta capacidad integradora permite a las plantas coordinar respuestas complejas de forma rápida y reversible, lo cual resulta esencial para su supervivencia y adaptación a entornos cambiantes.
Proteínas scaffold y su impacto en la funcionalidad de los SGs en plantas
La formación de los SGs no es un proceso aleatorio, sino que sigue una secuencia organizada de eventos. Estudios recientes proponen un modelo de tres etapas: primero, la nucleación de un núcleo denso (core) inicial; segundo, su crecimiento mediante el reclutamiento progresivo de ARN y proteínas; y finalmente, la formación de una región periférica más dinámica (shell), donde los componentes se intercambian continuamente con el citoplasma. Este modelo ayuda a entender cómo se distribuyen las funciones dentro del condensado. En las primeras etapas, las proteínas capaces de establecer múltiples interacciones, es decir, altamente multivalentes, desempeñan un papel clave al iniciar la formación del núcleo. Estas proteínas suelen actuar como scaffolds, proporcionando el andamiaje necesario para que el condensado se ensamble (Figura 2). Además de las proteínas, el ARN desempeña un papel activo como componente estructural, pudiendo funcionar como scaffold (Figura 2). Por ejemplo, la metilación m6A permite reclutar proteínas lectoras y aumentar su concentración local, facilitando la transición de fase al superar el umbral termodinámico necesario. Ejemplos como ECT1 y ECT8 en Arabidopsis ilustran cómo la interacción con ARN metilado regula la formación de condensados y modula la respuesta a estrés en plantas.
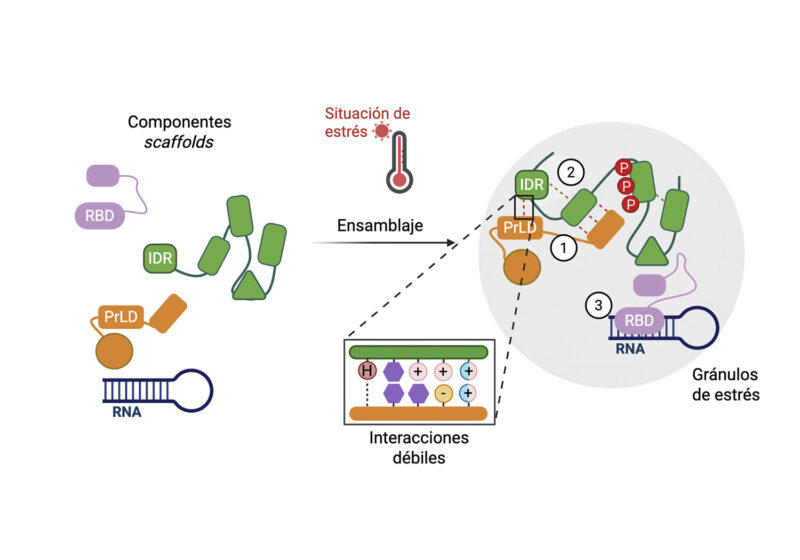
Mecanismos moleculares que impulsan el ensamblaje de los gránulos de estrés. La figura ilustra cómo, bajo una situación de estrés ambiental (p. ej. térmico, termómetro), los componentes scaffold (andamios) citoplasmáticos inician el proceso de auto-ensamblaje. Estos andamios moleculares incluyen proteínas multivalentes ricas en regiones intrínsecamente desordenadas (IDRs, por sus siglas en inglés), dominios tipo prión (PrLD), dominios de unión a ARN (RBD, por sus siglas en inglés) y moléculas de ARN. El ensamblaje del gránulo de estrés está impulsado termodinámicamente por una red cooperativa de interacciones débiles no covalentes. Se detallan tres tipos de interacciones clave: (1) interacciones hetero-típicas (IDR-PrLD) e (2) homo-típicas (IDR-IDR), las cuales pueden ser moduladas por modificaciones post-traduccionales como la fosforilación (marcadas con ‘P’), y (3) interacciones entre RBDs y ARN.
En el caso de las plantas, comienzan a identificarse proteínas que podrían desempeñar estos roles estructurales. Un ejemplo es la nucleasa Tudor Staphylococcal Nuclease (TSN, por sus siglas en inglés) que se acumula en los gránulos de estrés y está implicada en la regulación del ARN y la respuesta al estrés. Su capacidad para interactuar con múltiples componentes la convierte en una candidata clave para actuar como elemento organizador en estas primeras etapas del ensamblaje. De manera similar, otras proteínas de unión a ARN, como RBP47 y UBP1, han sido identificadas como componentes importantes que contribuyen a la nucleación y al ensamblaje de estos condensados. Se ha comprobado que la alteración de estas proteínas tiene un impacto severo en la composición, ensamblaje y dinámica de los SGs, así como en la respuesta vegetal al estrés. Por ejemplo, se ha descrito que la familia RBP47 actúa como un regulador negativo de la termotolerancia, ya que plantas mutantes knock-out (KO) para estos genes muestran una mayor tolerancia a condiciones de calor extremo, lo que sugiere que la correcta formación y dinámica de los SGs es determinante para la resiliencia y la supervivencia de las plantas en entornos adversos.
En conjunto, este marco permite conectar las propiedades físicas de los condensados con su función biológica: la multivalencia impulsa la nucleación, los scaffolds organizan la estructura y los clients aportan funcionalidad. Esta organización jerárquica da lugar a un sistema dinámico capaz de responder rápidamente a los desafíos del entorno.
Del sensor ambiental al diseño de cultivos: el paradigma de la separación de fases
Para comprender la formación de los SGs y en general de los condensados biomoleculares, es fundamental recurrir al concepto termodinámico de la separación de fases líquido-líquido (LLPS, por sus siglas en inglés liquid-liquid phase separation). De forma análoga a la mezcla de aceite y agua, ciertas macromoléculas celulares pueden agruparse espontáneamente en estructuras densas y dinámicas sin necesidad de una membrana. Este proceso permite que proteínas y ácidos nucleicos se concentren en regiones específicas, formando ensamblajes con propiedades reológicas similares a las de un líquido: capacidad de fusión, deformación e intercambio constante de componentes con el citoplasma.
A nivel molecular, la LLPS está impulsada por interacciones débiles y transitorias entre proteínas, especialmente en regiones intrínsecamente desordenadas (IDRs, por sus siglas en inglés intrinsically disordered proteins), que proporcionan flexibilidad y permiten múltiples contactos simultáneos (Figura 2). Entre estas interacciones destacan las electrostáticas, hidrofóbicas y, de forma clave, las interacciones π-π y catión-π, que contribuyen a la cohesión y dinámica de los condensados (Figura 2). Aunque este mecanismo es conservado en todos los organismos, en plantas las IDRs han adquirido funciones adicionales como sensores ambientales. Algunas proteínas utilizan estas regiones para percibir cambios físicos y desencadenar respuestas adaptativas. Por ejemplo, FUST1 actúa como termosensor promoviendo la formación de gránulos de estrés, SEUSS es clave en la respuesta al estrés osmótico, y ELF3 regula procesos de desarrollo en función de la temperatura mediante cambios en su estado de fase. En conjunto, estas propiedades permiten que las transiciones de fase traduzcan señales físicas del entorno en respuestas biológicas complejas a nivel celular y del organismo.
La LLPS no depende únicamente de la secuencia proteica, sino que está finamente regulada por modificaciones postraduccionales (PMTs, por sus siglas en inglés) que modulan la carga y multivalencia de las proteínas. En plantas, la fosforilación actúa como un interruptor clave que controla la formación de condensados en respuesta al estrés; por ejemplo, la proteína GRP7 regula su ensamblaje en función de su estado de fosforilación. Paralelamente, este mismo mecanismo orquesta procesos celulares vitales como la endocitosis mediada por clatrina; el estado de fosforilación de componentes del complejo TPLATE (como TASH3) modula la separación de fases de sus IDRs en la membrana plasmática. En conjunto, estos mecanismos ponen de manifiesto que la separación de fases no solo organiza la célula, sino que actúa como un sistema dinámico de percepción y respuesta al entorno. Comprender y manipular estos procesos abre nuevas oportunidades para la biotecnología vegetal, permitiendo diseñar cultivos capaces de adaptar de forma rápida y reversible su fisiología frente a condiciones ambientales cambiantes.
El reto metodológico: cómo descifrar los condensados en el sistema vegetal
El rápido crecimiento del campo de los condensados biomoleculares ha ido acompañado de una gran diversidad de enfoques experimentales y computacionales. Sin embargo, en plantas, el estudio de estos ensamblajes sigue siendo metodológicamente heterogéneo. Muchas de las herramientas utilizadas proceden de sistemas animales, y no siempre se adaptan bien a las particularidades de las células vegetales, como la presencia de una pared celular rígida, la presión de turgencia o la autofluorescencia de los cloroplastos.
En general, el estudio de los condensados combina tres niveles de aproximación: análisis computacionales, experimentos in vitro y observaciones en células vivas (Figura 3). Los enfoques in silico suelen ser el punto de partida, ya que permiten identificar proteínas con características que favorecen la formación de condensados, como regiones intrínsecamente desordenadas o motivos que facilitan interacciones múltiples. No obstante, estas predicciones deben interpretarse con cautela, especialmente en plantas, donde muchas proteínas presentan señales de direccionamiento a orgánulos o regiones de baja complejidad que pueden complicar el análisis. Como ejemplo concreto en este nivel, la comunidad investigadora recurre actualmente a bases de datos curadas como PhaSepDB o CD-CODE, y a predictores avanzados basados en aprendizaje profundo (deep learning) como ProtGPS; este último utiliza modelos de lenguaje de proteínas (ESM2) para predecir no solo la propensión a la separación de fases, sino también la localización de una proteína en condensados específicos (Figura 3, panel izquierdo).
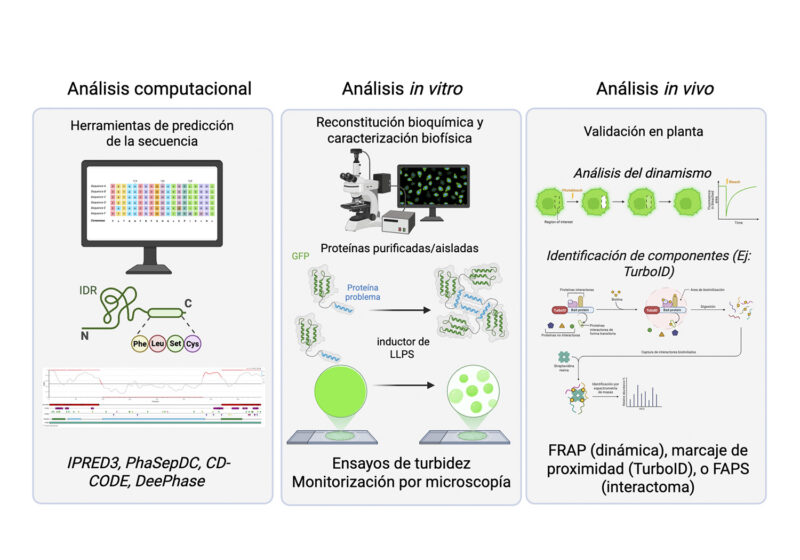
Enfoque multidisciplinar para el estudio de condensados en plantas. El análisis se divide en tres frentes complementarios: (1) Análisis computacional (in silico) para la predicción de proteínas con capacidad de separación de fases (LLPS) mediante modelos bioinformáticos; (2) Análisis in vitro para validar la formación y morfología de las gotas líquidas mediante ensayos de turbidez y microscopía con proteínas purificadas; y (3) Análisis in vivo para caracterizar sus propiedades fisiológicas, utilizando FRAP para medir el dinamismo del condensado y técnicas de biotinilación por proximidad (p. ej. TurboID) para identificar las redes de interacción proteica dentro de la célula viva.
Los ensayos de reconstitución in vitro continúan siendo el estándar más empleado para evaluar la capacidad intrínseca de una proteína a experimentar LLPS (Figura 3, panel central). El procedimiento clásico requiere la expresión heteróloga de la proteína de interés, generalmente fusionada a un marcador fluorescente, en sistemas bacterianos como E. coli, seguida de su purificación. Posteriormente, la transición de fase se induce in vitro mediante la alteración sistemática de parámetros fisico-químicos del tampón, tales como la fuerza iónica, el pH o la temperatura (inductor de LLPS). La formación de condensados se monitoriza cuantitativamente mediante ensayos de turbidez y se caracteriza a nivel morfológico mediante microscopía, documentando la nucleación, morfología esférica y coalescencia de las gotas líquidas. Sin embargo, dado que estas condiciones diluidas idealizadas no reflejan la alta densidad macromolecular de una célula viva, el campo está experimentando una transición metodológica. En la actualidad, resulta crítico complementar estos ensayos básicos mimetizando el entorno fisiológico vegetal; esto se logra incorporando agentes de hacinamiento molecular (molecular crowding, como el PEG o el dextrano) y modulando el estado redox del medio, lo que permite evaluar la termodinámica del condensado bajo condiciones que aproximan la complejidad del citoplasma.
Demostrar la relevancia fisiológica de los condensados exige su caracterización in vivo, un reto complejo en plantas debido a interferencias ópticas (como la autofluorescencia) y a la necesidad de un control de expresión riguroso sin sobreexpresión. Su estudio actual requiere enfoques multidisciplinares divididos en dos frentes. Por un lado, para definir sus propiedades biofísicas, y distinguir un estado líquido funcional de un agregado irreversible, técnicas clásicas como el FRAP se combinan con micro-reología o sondas específicas (como la Tioflavina-T). Por otro lado, para mapear su composición, dadas sus interacciones débiles y transitorias, se emplean estrategias de aislamiento avanzado (como la clasificación de partículas FAPS) y mapeo in situ del interactoma mediante marcaje por proximidad, destacando especialmente el uso de enzimas ligasas como TurboID (biotinilación in vivo), así como técnicas de entrecruzamiento químico (crosslinking).
Retos y perspectivas
El estudio de los condensados biomoleculares en plantas enfrenta barreras biofísicas y ópticas únicas, como la pared celular rígida, las grandes vacuolas y la autofluorescencia de los cloroplastos, lo que exige una urgente estandarización metodológica para evitar artefactos experimentales. El gran reto actual es capturar estas interacciones dinámicas en su entorno celular real para desentrañar cómo las proteínas scaffold, los elementos client, el ARN y las modificaciones químicas orquestan esta compleja respuesta. Superar estas limitaciones técnicas y conceptuales permitirá comprender en profundidad cómo las plantas pueden utilizar la LLPS como un sensor ambiental directo, un conocimiento básico indispensable para desarrollar, en el futuro, estrategias que mejoren la resiliencia agrícola frente al cambio global.
Agradecimientos
Este trabajo se ha desarrollado en el grupo del Dr. Emilio Gutiérrez-Beltrán denominado «Gránulos de estrés y señalización» y se encuentra financiado por los proyectos PID2023-152750NB-I00 (MCIN/ AEI /10.13039/501100011033/FEDER, UE) y DGP_PIDI_2024_02706 (Junta de Andalucía).
Para leer más
- Gutiérrez-Beltrán E, et al. Tudor staphylococcal nuclease is a docking platform for stress granule components and is essential for SnRK1 activation in Arabidopsis. EMBO Journal 40 (2021) e105043. https://doi.org/10.15252/embj.2020105043
- Liu X, Zhu JK, Zhao C. Liquid-liquid phase separation as a major mechanism of plant abiotic stress sensing and responses. Stress Biology 3 (2023) 56. https://doi.org/10.1007/s44154-023-00141-x
- Peng J, Yu Y, Fang X. Stress sensing and response through biomolecular condensates in plants. Plant Communications 6 (2025) 101225. https://doi.org/10.1016/j.xplc.2024.101225
- Solis-Miranda J, et al. Stress-related biomolecular condensates in plants. The Plant Cell 35 (2023) 3187–3204. https://doi.org/10.1093/plcell/koad127
- Xie Z, et al. Phenolic acid-induced phase separation and translation inhibition mediate plant interspecific competition. Nature Plants 9 (2023) 1481-1499. https://doi.org/10.1038/s41477-023-01499-6
