
Durante décadas, el estudio del trastorno del espectro autista (TEA) ha estado dominado por la genética: mutaciones, variantes raras y redes de genes implicados en el neurodesarrollo. Sin embargo, una línea de investigación emergente está desplazando parcialmente el foco hacia un nivel más sutil, y profundamente fascinante, de organización biológica: las propiedades físicas de ensamblajes de proteínas dentro de la célula. En este contexto, la proteína CPEB4 (por sus siglas en inglés, Cytoplasmic Polyadenylation Element Binding protein 4) ha pasado de ser un regulador más de la expresión génica a convertirse en un actor relevante en el TEA idiopático (no sindrómico), especialmente a través de su capacidad de formar condensados y agregados.
CPEB4: un regulador maestro de la traducción
CPEB4 pertenece a una familia de proteínas que controlan la traducción de ARN mensajeros (ARNm) mediante la regulación de la longitud de sus colas de poli-A, un mecanismo clave para determinar si un ARNm permanece silenciado o es activamente traducido a proteína. En concreto, CPEB4 se une a secuencias específicas (llamadas CPE) en los ARNm diana y, en función de su estado y del contexto celular, puede reclutar complejos que alargan la cola de poli-A, favoreciendo la iniciación de la traducción, o bien mantener estos transcritos en un estado reprimido.
Este control es especialmente relevante en neuronas, donde la traducción localizada y temporalmente precisa resulta esencial para el desarrollo y la plasticidad sináptica. Más que actuar sobre genes aislados, CPEB4 funciona como un regulador maestro que coordina la traducción de múltiples ARNm. Entre sus dianas se encuentran numerosos genes asociados al riesgo de TEA, lo que sugiere que alteraciones en la actividad de CPEB4 pueden tener efectos sobre la función neuronal.
En 2018, se vinculó por primera vez directamente CPEB4 con el autismo idiopático. En muestras de cerebros humanos post mortem se observó una desregulación tanto de la poliadenilación como de los niveles de múltiples ARNm controlados por CPEB4. De forma llamativa, esta alteración no se debía a mutaciones en el gen CPEB4, sino a cambios en su procesamiento. CPEB4 presenta varias isoformas generadas por splicing alternativo, entre las cuales destaca la inclusión o exclusión de un pequeño exón, el microexón 4 (24 nucleótidos, 8 aminoácidos), que representa apenas un 1% de la secuencia total de la proteína. Tanto las isoformas que incluyen este microexón como las que carecen de él se expresan de manera natural en neuronas, en proporciones finamente definidas. Sin embargo, en el TEA idiopático se ha descrito un cambio de splicing que altera este equilibrio, favoreciendo la producción de la isoforma que carece del microexón (Figura 1). Este cambio, aunque sutil, tiene consecuencias importantes sobre las propiedades funcionales de la proteína.
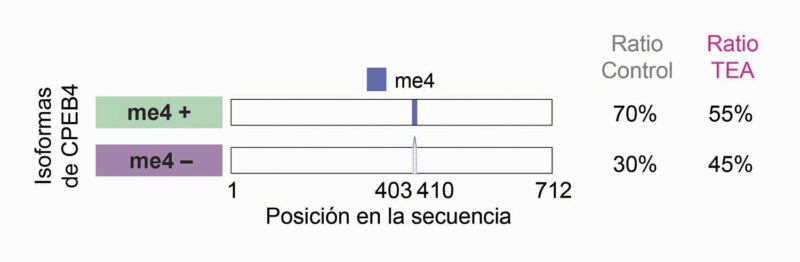
Representación de las isoformas de CPEB4. Representación de dos isoformas de CPEB4, conteniendo (me4+) o no (me4-) el microexón neuronal (me4, azul). El promedio de las ratios de las isoformas en el control o TEA idiopático se indica a la derecha.
Condensación biomolecular: cuando las proteínas forman «gotas»
Muchas proteínas con regiones intrínsecamente desordenadas —como CPEB4, cuyo dominio desordenado abarca más de la mitad de su longitud total— pueden organizarse en condensados dinámicos, estructuras similares a «gotas líquidas» que concentran moléculas específicas sin necesidad de membrana. Estos condensados contienen centenares de moléculas de CPEB4, junto con otras proteínas y ARN, creando microambientes funcionales dentro de la célula. En condiciones fisiológicas, los condensados de CPEB4 son altamente reversibles, lo que permite una regulación espacial y temporal extremadamente precisa del proceso de la traducción.
La dinámica de ensamblaje y disolución de los condensados es un proceso que está finamente regulado por señales celulares específicas. En células en ciclo celular, la forma no fosforilada de la proteína tiende a ensamblarse en condensados, en cuyo caso actúa como represor de la traducción, en fases no mitóticas. En cambio, en meiosis y mitosis, la hiperfosforilación de CPEB4 en su dominio desordenado mediada por las quinasas Cdk1 y ERK2 promueve la disolución de los condensados y mantiene a la proteína en un estado monomérico y activo. La adición de grupos fosfato incrementa la carga neta de la proteína, reduciendo su capacidad de asociarse y favoreciendo la dispersión de los condensados en momentos concretos.
En neuronas, sin embargo, el mecanismo que controla la formación/disolución de los condensados de CPEB4 es distinto. En este caso, la disolución de los condensados de CPEB4 está vinculada a cambios transitorios en el pH intracelular que ocurren durante la estimulación neuronal. El dominio desordenado de CPEB4 está enriquecido en residuos de histidina, un aminoácido cuya carga depende de manera muy sensible del pH. Los condensados de CPEB4 están estabilizados por interacciones mediadas por histidinas. Así, cambios sutiles entorno al pH fisiológico, como ocurre durante la estimulación neuronal, cambian la carga de las histidinas y consecuentemente su capacidad de formar interacciones, afectando a la estabilidad de los condensados (Figura 2).
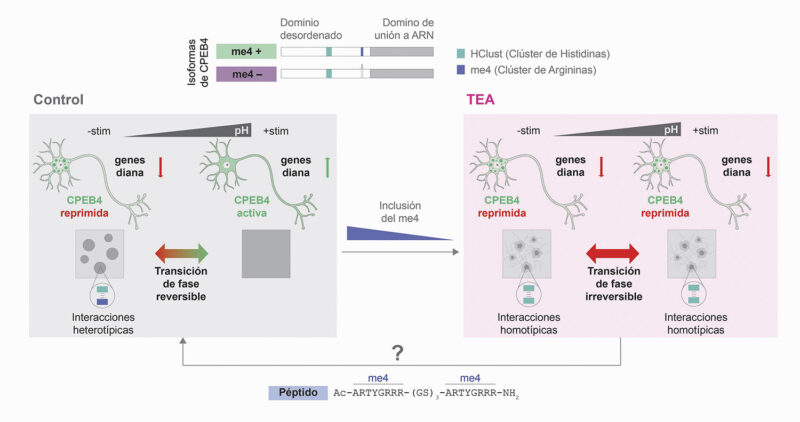
Resumen esquemático del proceso de la regulación de la traducción de CPEB4 mediante la formación de condensados biomoleculares. En el panel de la izquierda se ilustra el proceso de condensación reversible de CPEB4 en condiciones fisiológicas, donde los condensados dinámicos de disuelven durante la estimulación neuronal (+stim) debido a cambios de pH intracelulares. En el panel de la derecha se ilustra el proceso de agregación de CPEB4 en ausencia del microexón, que impide la disolución de los condensados y como consecuencia no se promueve la traducción de genes diana. Adaptado de Garcia-Cabau, Bartomeu, et al. Nature (2025).
De la condensación a la agregación: el hallazgo clave
Sin embargo, estos condensados no son siempre estables. Bajo ciertas condiciones, pueden transicionar hacia estados menos dinámicos, más sólidos, o incluso agregados. Este tipo de transiciones ha cobrado gran relevancia en biomedicina, ya que se ha observado en diversas enfermedades neurodegenerativas, como el Alzheimer (con la proteína tau) o la ELA (con las proteínas TDP-43 y FUS), donde proteínas inicialmente funcionales que forman condensados biomoleculares acaban formando agregados patológicos. A diferencia de los condensados líquidos, que son dinámicos y reversibles, los agregados suelen ser irreversibles. En el caso de CPEB4, la diferencia de reversibilidad que hay entre estos dos estados resulta crítica, ya que su función depende de la capacidad de los condensados de ensamblarse y disolverse en respuesta a la estimulación neuronal.
En 2025, se demostró que CPEB4 puede sufrir una transición de los condensados funcionales a agregados sólidos en ausencia del microexón 4. Experimentos in vitro con proteína purificada mostraron que los condensados pierden su carácter dinámico y evolucionan hacia estructuras fibrilares, lo que implica la pérdida de su reversibilidad, es decir, de su capacidad para disolverse. Estas observaciones se confirmaron posteriormente en sistemas celulares, donde la isoforma sin microexón mostraba una menor capacidad de disolución comparado con la isoforma que contiene el microexón. Finalmente, un modelo de ratón que reproduce las proporciones de isoformas observadas en el autismo idiopático también evidenció la presencia de agregados de CPEB4 en cortes de cerebro, reforzando la relevancia fisiopatológica de este fenómeno (Figura 2).
¿Cómo funciona el efecto protector del microexón 4 frente a la agregación de CPEB4? Este microexón introduce residuos con carga positiva (argininas) que pueden establecer interacciones con histidinas de la secuencia. Las regiones ricas en histidinas presentes en la propia proteína pueden interaccionar de forma homotípica (histidina-histidina) y dar lugar a la formación de agregados. La presencia del microexón parece neutralizar parcialmente este efecto al competir por estas interacciones, generando interacciones heterotípicas (histidina-arginina) y reorganizando la red de contactos, reduciendo la probabilidad de que las histidinas participen en asociaciones homotípicas. De este modo, el microexón actúa como un elemento atenuador que ajusta el balance de interacciones intermoleculares en los condensados de CPEB4, limitando su tendencia a formar agregados (Figura 2).
Si la disfunción de CPEB4 se debe a un cambio en las propiedades físicas de sus condensados, surge una pregunta inevitable: ¿es posible revertirlo? ¿se pueden recuperar las propiedades dinámicas de los condensados? Resultados preliminares sugieren que sería posible. Un péptido sintético basado en la secuencia del microexón 4 es capaz de prevenir la agregación de la isoforma de CPEB4 que carece de este fragmento, restaurando in vitro la dinámica y reversibilidad de los condensados. Aunque estas aproximaciones se encuentran todavía en fases iniciales, abren una vía terapéutica novedosa basada en la modulación de las propiedades físicas de los condensados proteicos (Figura 2).
Conclusión
La historia de CPEB4 en el autismo idiopático ilustra cómo la biología está integrando conceptos de la física para comprender tanto la fisiología como la patología. Lo que comenzó como una alteración en la regulación de la traducción ha evolucionado hacia un modelo en el que la condensación y agregación proteica ocupan un lugar central. Este nuevo marco es cada vez más común y proporciona explicaciones coherentes para observaciones previas en distintos sistemas proteicos. También redefine las posibilidades terapéuticas, sugiriendo que intervenir sobre las propiedades físicas de los condensados proteicos podría ser una estrategia válida en patologías donde se producen transiciones aberrantes de estados líquidos a sólidos.
Para leer más
- Conicella A, Zerze GH, Mittal J, Fawz NL. ALS Mutations Disrupt Phase Separation Mediated by α-Helical Structure in the TDP-43 Low-Complexity C-Terminal Domain. Structure 24 (2016) 1537-1549. https://doi.org/10.1016/j.str.2016.07.007
- Garcia-Cabau C, Bartomeu A, et al. Mis-splicing of a neuronal microexon promotes CPEB4 aggregation in ASD. Nature 637 (2025) 496–503. https://doi.org/10.1038/s41586-024-08289-w
- Guillén-Boixet J, Buzon V, Salvatella X, Méndez R. CPEB4 is regulated during cell cycle by ERK2/Cdk1-mediated phosphorylation and its assembly into liquid-like droplets. eLife 5 (2016) e19298. https://doi.org/10.7554/eLife.19298
- Parras A, Anta H, Santos-Galindo M, et al. Autism-like phenotype and risk gene mRNA deadenylation by CPEB4 mis-splicing. Nature 560 (2018) 441–446. https://doi.org/10.1038/s41586-018-0423-5
- Patel A, Lee H O, Jawerth L, et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell 162 (2015) 1066-1077. https://doi.org/10.1016/j.cell.2015.07.047
- Wegmann S, Eftekharzadeh B, Tepper K, et al. Tau protein liquid–liquid phase separation can initiate tau aggregation. EMBO Journal 37 (2018) e98049. https://doi.org/10.15252/embj.201798049
