
El título de este artículo puede traer a la mente recuerdos escolares de los intercambios de fase entre el hielo, el agua y el vapor del agua. En la primera parte de este ensayo, revisaremos algunos de estos intercambios «sencillos» en procesos fisiopatológicos de gran importancia (Figura 1). La segunda parte del ensayo tratará de otra clase de intercambio de fases, las separaciones de fase líquido/líquido (LLPS, por sus siglas en inglés Liquid-Liquid Phase Separation). La separación de fase líquido/líquido ha ganado gran relevancia durante los últimos 17 años en la biología molecular y celular por su capacidad de regular y fomentar procesos bioquímicos (Figura 2).
Quizás resulte sorprendente que el primer cambio de fases que aprendimos en el cole, la transición entre agua y hielo, esté modulado de forma compleja por la biología. El hielo se funde a 0ºC, pero el agua muy pura puede superenfriarse hasta -40ºC sin congelarse ya que el núcleo de hielo contiene al menos 90 moléculas de agua y la entropía se opone fuertemente a su formación. Organismos que habitan en climas fríos cuentan con péptidos y/o proteínas anticongelantes que atrapan los núcleos nacientes de hielo o los obligan a crecer distorsionadamente, retrasando su desarrollo. Estas proteínas anticongelantes tienen orígenes evolutivos diversos. Sus estructuras también son diferentes aunque comparten una cara plana que se une al hielo. Una de estas proteínas tiene una estructura fascinante: está compuesta en casi su 50% por residuos de glicina y consiste en un haz de seis hélices de poliprolina II (PPII) organizada en una bicapa (Figura 1).
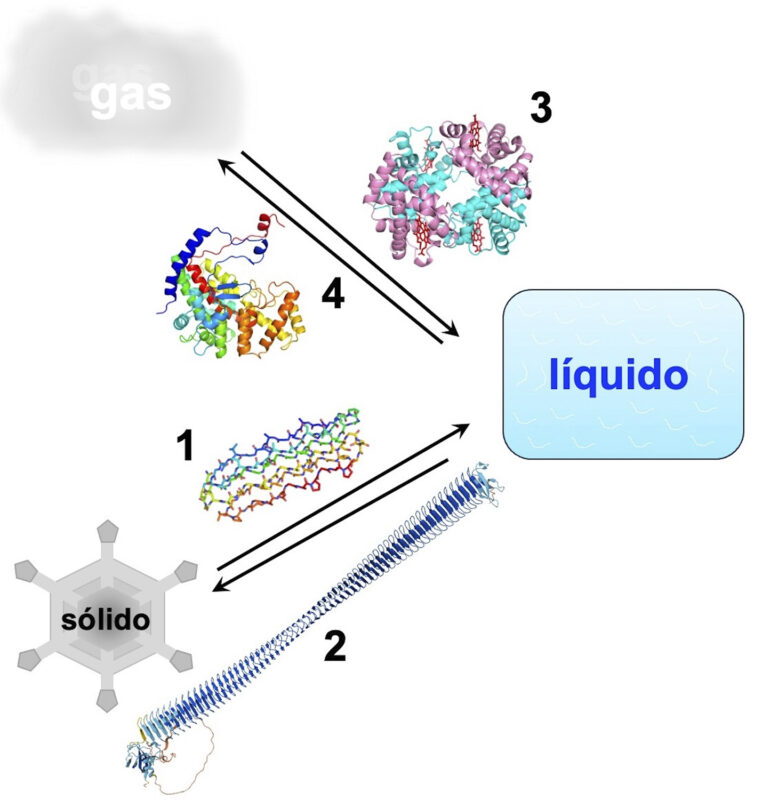
Intervención de proteínas (1) anticongelantes, (2) inductores de congelación ELSA (por sus siglas en inglés, enhanced ligand for solid aqua), (3) hemoglobina y (4) anhidratasa carbónica en transiciones entre las fases sólida, líquida y gaseosa.
De la misma forma que hay proteínas que inhiben el cambio de fase del agua al hielo, hay otras que lo promueven de tal forma que van ordenando las moléculas del agua en estructuras similares al núcleo de hielo (Figura 1). Así logran que el agua se congele a temperaturas ligeramente inferiores a 0 ºC. Un mejor ejemplo de esto se encuentra en la bacteria Pseudomonas syringae, que induce la formación de hielo para dañar la pared celular en frutas, creando una entrada para la bacteria. Aunque P. syringae estropea la fruta, también juega un papel beneficioso en la producción de nieve artificial en las estaciones de esquí. Más relevante aún es su papel ecológico en el ciclo del agua, ya que grandes cantidades de esta bacteria son transportadas por el viento y una fracción significativa de la nieve y la lluvia que cae sobre la faz de la Tierra es inducida por P. syringae y microbios similares.
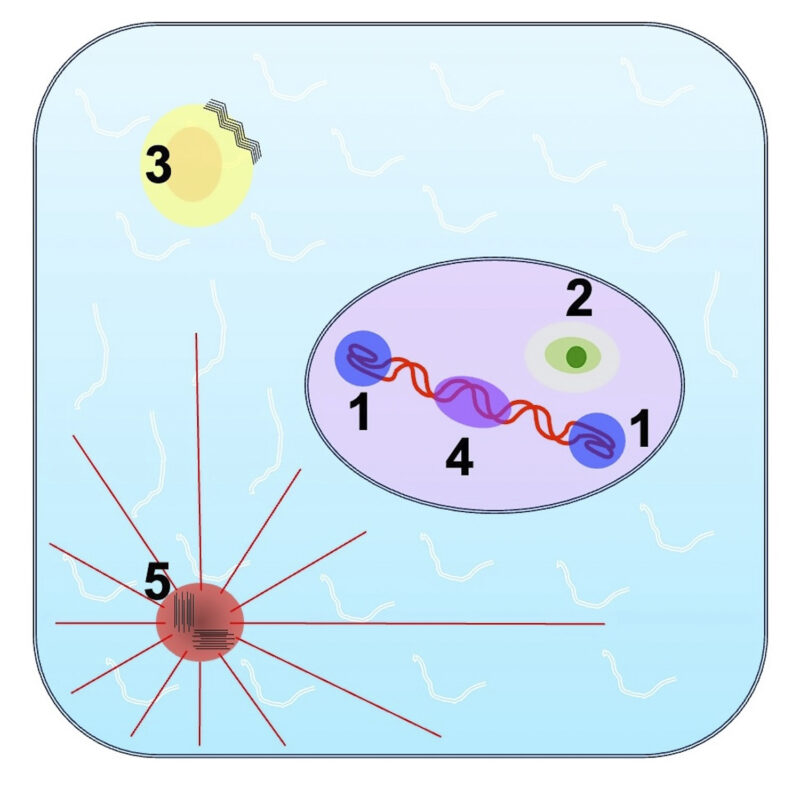
Algunos condensados biomoleculares: (1) el cuerpo de Cajal, (2) el nucleolo, (3) el gránulo de estrés, que puede promover la formación de amiloides (líneas onduladas), (4) el complejo potenciador y (5) la matriz pericentriolar.
Cuando respiramos, las moléculas de oxígeno en fase gaseosa pasan a su fase sólida en combinación con la proteína hemoglobina y moléculas de dióxido de carbono, bien unidas en fase sólida a hemoglobina o disueltas en fase líquida como CO2 o H2CO3, y son liberadas a la atmósfera en fase gaseosa (Figura 1). Estos fueron los primeros procesos fisiológicos dilucidados a nivel molecular gracias a los estudios de Perutz, Kendrew y sus colaboradores. Aunque menos conocidos que la hemoglobina, las proteínas y lípidos que componen el surfactante pulmonar son igualmente importantes para estos cambios de fase. La tensión superficial agua/aire es tan elevada que los insectos la aprovechan para andar sobre el agua y los pulmones de bebés prematuros no la superan para respirar. El surfactante pulmonar reduce esta tensión superficial y gracias a continuas investigaciones sobre su composición y función ha sido posible desarrollar preparaciones para salvar bebés prematuros.
Nuestro conocimiento sobre el estado de fase de las células ha ido evolucionando a lo largo de los últimos 125 años. A principios del siglo XX, gracias a estudios de microscopía de luz visible se había establecido la existencia de varios organelos «sólidos» como la mitocondria, el núcleo, el aparato de Golgi, etc. que flotan en un citoplasma de carácter líquido viscoso (Figura 3). Entonces, el interior de la célula podría describirse como un coloide, es decir, una mezcla de un líquido y partículas sólidas insolubles minúsculas (de 1 a 1.000 nm), que dispersan la luz y no se asientan con el tiempo. También se reportaron otros organelos, como el nucleolo, el cuerpo de Cajal, o el centrosoma cuya presencia era menos constante y depende de las condiciones celulares.
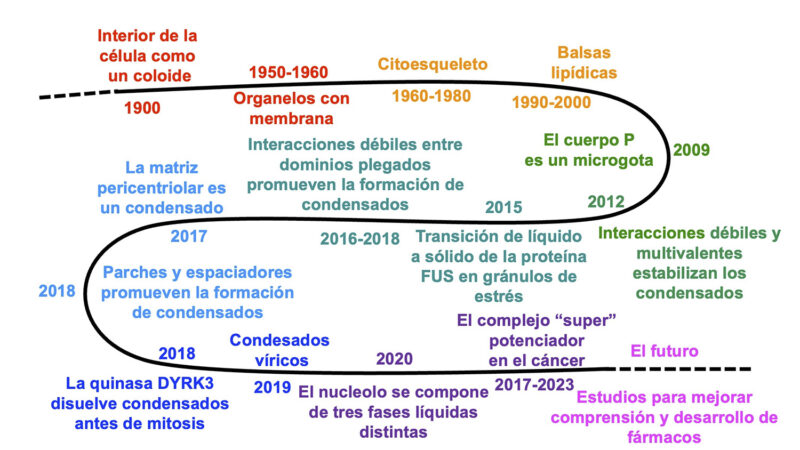
Cronograma de descubrimientos principales en transiciones de fase intracelulares.
El siguiente gran avance surgió en los años 1950-1960. Gracias a mejoras en microscopía electrónica, se descubrió que muchos de los organelos principales, como el núcleo, la mitocondria, el retículo endoplasmático, el aparato de Golgi o las vesículas están rodeados por membranas. Esto dio lugar a un nuevo modelo celular con las membranas como las protagonistas de la organización del interior de la célula. No obstante, la estructura de los organelos sin membrana como el nucleolo, el cuerpo de Cajal o el centrosoma siguió siendo un misterio. Por su parte, la visión del citoplasma cambió en las décadas de 1960-1980 con descubrimientos que dotaron a ese líquido viscoso un citoesqueleto de carácter dinámico compuesto por microtúbulos, filamentos intermedios y filamentos de actina.
Este viaje platónico de la pegajosidad caótica hacia la estructura y el orden promovido por membranas y filamentos dio un giro inesperado en la década de 1990-2000 con los descubrimientos relativos a la membrana celular que indicaban que podría organizarse en zonas menos fluidas con una composición molecular distinta llamada «balsa lipídica». Este cambio de fase líquido/líquido en el contexto de la membrana tiene funciones importantes en la transmisión de señales y presagió el descubrimiento de la naturaleza real de los organelos sin membrana.
Un avance verdaderamente revolucionario ocurrió en 2009 de la mano de Brangwynne, Hyman y sus colaboradores. Ellos descubrieron que un organelo especial llamado cuerpo P (o P-body), el cual determina el destino de la línea de células germinales durante el desarrollo, se comporta como una microgota de líquido separada por fases. Para estudiar los cuerpos P, emplearon una técnica denominada FRAP (recuperación de la fluorescencia tras el fotoblanqueo, por sus siglas en inglés). En ella, se utiliza un láser para decolorar (fotoblanquear) la mitad de un cuerpo P marcado con proteínas fluorescentes. Al medir cuánto tardan las moléculas de la mitad no blanqueada en mezclarse con la parte afectada, pudieron calcular la viscosidad del cuerpo P, descubriendo que es similar a la de líquidos viscosos como la miel.
En otros experimentos, se fotoblanqueaba la gota completa. Al medir la recuperación de la fluorescencia en el orgánulo totalmente decolorado, lograron calcular la tasa de intercambio de moléculas entre el cuerpo P y el resto del citoplasma.
Estos descubrimientos impulsaron el estudio de otros orgánulos sin membrana o «condensados biomoleculares», como se les denomina habitualmente hoy en día. En los últimos 17 años, se ha informado de la existencia de más de 24 condensados formados por separación de fases líquido-líquido (LLPS). Muchos están implicados en la síntesis, modificación, corte y empalme, y regulación de traducción del ARN. Entre los más notables se encuentran: 1. El cuerpo de Cajal: clave en la acción de la telomerasa; 2. El nucleolo: que funciona como una línea de ensamblaje para los ribosomas y contiene tres capas distintas separadas por fases; 3. El gránulo de estrés: se forma de manera transitoria ante condiciones celulares adversas para detener la traducción, reteniendo ARNm y ciertas proteínas; 4. El complejo potenciador (Enhancer): se forma alrededor del dominio C-terminal de la ARN polimerasa II y sirve para potenciar drásticamente la transcripción; y 5. La matriz pericentriolar que rodea a los centriolos y organiza la red de microtúbulos del citoesqueleto (Figura 2).
Al estudiar el comportamiento de proteínas y ARNs marcados bajo diversas condiciones (como el estrés o la mitosis) y observar los efectos de las mutaciones en la LLPS, los científicos establecieron que ciertas moléculas, llamadas «andamios», son esenciales para la formación del condensado. Otras macromoléculas, denominadas «clientes», solo pueden integrarse una vez que el condensado ya se ha formado. Asimismo, se descubrió que las interacciones débiles, transitorias y multivalentes son fundamentales para mantener el condensado formado en fase líquido. Estas interacciones pueden estar mediadas por: 1. Homo-oligomerización de estructuras plegadas: como ocurre con los dominios N-terminales de la TDP-43 en los gránulos de estrés y de la nucleoplasmina en el nucleolo; 2. Segmentos ricos en prolina: que se unen débilmente a una serie de dominios SH3. 3. Residuos «pegajosos», es decir D, E, F, I, K, L, M, R, W e Y, presentes en regiones intrínsecamente desordenadas (IDRs, por sus siglas en inglés intrinsically disordered regions), que median interacciones electrostáticas (catión/anión), catión/π, π/π e hidrofóbas.
Los residuos pegajosos suelen estar dispersos entre residuos «espaciadores» flexibles como la alanina (A), serina (S) y treonina (T). Aunque la glicina (G) también se considera un espaciador flexible, los segmentos con varias glicinas consecutivas o repeticiones (GGX)N pueden ensamblarse en haces de hélices de PPII. Por último, los residuos de S y T son cruciales debido a su capacidad de ser fosforilados; de hecho, la fosforilación es un mecanismo común que provoca la disolución de los condensados. Por ejemplo, al iniciar la mitosis, la acción de la quinasa DYRK3 lleva a la disolución de los gránulos de estrés, motas nucleares de empalme y la matriz pericentriolar. Más tarde, esta enzima es degradada al terminar la mitosis, lo cual facilita la reconstitución de dichos condensados. Otras clases de modificación, por ejemplo la oxidación y reducción de cisteína, la glicosilación, acetilación, ubiquitinación, metilación, etc. también pueden modular el equilibrio entre la fase dispersa y la condensada.
Si bien estos experimentos proporcionaron conocimientos fundamentales, es posible que la presencia de marcadores fluorescentes o proteínas de fusión puedan inducir perturbaciones en el sistema. Asimismo, dichos estudios no ofrecen información a resolución atómica sobre las preferencias conformacionales, la dinámica o las interacciones moleculares. Otras técnicas, como la dispersión de rayos X de bajo ángulo (SAXS, por sus siglas en inglés), las simulaciones de dinámica molecular (DM) y la espectroscopia de resonancia magnética nuclear (RMN), han permitido subsanar estas carencias.
La técnica SAXS ha aportado datos clave sobre la nucleación de los condensados. Por su parte, las simulaciones de DM han validado el modelo de ‘stickers and spacers’ (parches y espaciadores) y han demostrado que la liberación de moléculas de agua del condensado favorece su formación desde un punto de vista entrópico. Además, han evidenciado que los grupos de residuos cargados u otros ‘stickers’ promueven la asociación con mayor eficacia cuando están agrupados que cuando se encuentran dispersos en el entorno. Finalmente, estas simulaciones sugieren que la interfaz entre el condensado y el medio circundante presenta una alta densidad de grupos con valencias no satisfechas, lo que tiende a favorecer diversas reacciones y asociaciones. Por otro lado, la espectroscopia de RMN ha suplido las limitaciones de la DM (campos de fuerza y tiempo computacional) que limita el estudio de sistemas tan complejos, lo que ha permitido revelar que las proteínas en estado condensado presentan una menor movilidad que en la fase dispersa, aunque mantienen un grado de desorden similar. En ciertos casos, se ha observado un incremento parcial en la formación de estructuras de hélice alfa o lámina beta dentro de la fase condensada.
Estos estudios también han demostrado que la viscosidad de los condensados es variable. Aquellos que se forman y disuelven según las condiciones celulares, como los gránulos de estrés, suelen tener viscosidades bajas. Por el contrario, la matriz pericentriolar, que debe resistir tensiones mecánicas, es muy viscosa y presenta una consistencia similar a la de un gel. Algunos condensados, como el cuerpo de Balbiani, experimentan una transición de fase completa de líquido a sólido para inducir cambios a largo plazo en el metabolismo celular. La viscosidad de estos condensados se regula mediante la fosforilación, así como por la acción de chaperonas y ARN helicasas. Cuando esta regulación falla, los condensados pueden endurecerse, lo que favorece el cambio de fase de proteínas como TDP-43 y FUS en amiloides, estos procesos han sido vinculados a enfermedades neurodegenerativas como la Esclerosis Lateral Amiotrófica (ELA). Dado que numerosos condensados desempeñan funciones esenciales en la bioquímica del ARN, no es de extrañar que su disfunción contribuya a patologías como el síndrome de Rett o la atrofia muscular espinal. En la última, la proteína de supervivencia de las neuronas motoras no logra separarse de fase correctamente, lo que provoca un ensamblaje defectuoso de los espliceosomas.
Al igual que las bacterias pueden manipular la transición de fase del agua a hielo, los virus y las células cancerosas pueden secuestrar de forma análoga la LLPS y subvertir los condensados biomoleculares para favorecer su propio crecimiento. Algunos virus inducen la formación de nuevos condensados para optimizar la producción de ARN y proteínas virales, el ensamblaje de nuevos viriones, o ambos. También manipulan los condensados existentes para secuestrar factores de defensa de la célula huésped. Asimismo, los virus suelen inducir la formación de gránulos de estrés para bloquear la síntesis de proteínas de la célula infectada, liberando así recursos para la producción de componentes virales. En ciertas leucemias, la proteína del poro nuclear Nup98 es secuestrada hacia el núcleo y combinada con la proteína Hoxa9 para fortalecer el complejo potenciador (enhancer) y «turbocargar» la transcripción. Por otro lado, en la mayoría de los cánceres, los nucléolos aumentan en tamaño y número y producen demasiados ribosomas. Esto se debe a mutaciones en el gen Myc, que activa la ARN polimerasa I, disparando la síntesis de ARNr.
Esta nueva comprensión de las funciones, mecanismos y patologías de los cambios de fase subcelulares ha impulsado el desarrollo de tratamientos innovadores. Actualmente se están probando diversos enfoques como moduladores de fluidez como la lipoamida que aumentan la fluidez de los gránulos de estrés para intentar bloquear las transiciones de fase patológicas de proteínas como TDP-43 o FUS. Por otra parte, ligandos quiméricos pueden dirigir proteínas específicas relacionadas con la disfunción de condensados hacia el proteasoma para su degradación. Otros compuestos buscan disolver los condensados virales o forzar una transición de fase de líquido a sólido para inutilizarlos. Por ejemplo, se ha demostrado que el antibiótico kanamicina reduce la LLPS y bloquea la formación de condensados del SARS-CoV-2, deteniendo la replicación del virus. Recientemente se ha reportado un nuevo mecanismo para otro fármaco antiguo, el cisplatino. Aunque durante mucho tiempo se creyó que este potente antitumoral actuaba únicamente mediante el entrecruzamiento del ADN, se ha descubierto que se concentra selectivamente en los condensados de los potenciadores en el núcleo, multiplicando su eficacia. Paralelamente, en el citoplasma, el cisplatino impulsa la formación irreversible de gránulos de estrés modificados, lo que provoca errores letales en la traducción.
Dada la relevancia fundamental y la importancia médica de las transiciones de fase en biología, este campo seguirá captando el interés de investigadores, la industria farmacéutica y la medicina en los próximos lustros.
Para leer más
- Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Jülicher F, Hyman AA. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 324 (2009) 1729-32. https://doi.org/10.1126/science.1172046
- Christner BC, Morris CE, Foreman CM, Cai R, Sands DC. Ubiquity of biological ice nucleators in snowfall. Science 319 (2008) 1214. https://doi.org/10.1126/science.1149757
- Collada A, Cruz A, Pérez-Gil J. Studying the interfacial activity and structure of pulmonary surfactant complexes. Chemistry and Physics of Lipids 266 (2025) 105459. https://doi.org/10.1016/j.chemphyslip.2024.105459
- Rai AK, Chen JX, Selbach M, Pelkmans L. Kinase-controlled phase transitions of membraneless organelles in mitosis. Nature 559 (2018) 211-216. https://doi.org/10.1038/s41589-018-0279-8
