Las plantas tienen que ajustar continuamente su fisiología y crecimiento a condiciones medioambientales cambiantes. Dicho ajuste implica intrincadas redes de señalización cuyo efecto final es la activación o represión de genes necesarios para adaptarse a los cambios en el ambiente. En estos procesos de adaptación desempeñan un papel crucial cambios moleculares reversibles en la cromatina, constituida por el enrollamiento del ADN alrededor de proteínas denominadas histonas. Tales cambios, que incluyen diversas modificaciones químicas de las histonas y el ADN, reciben el nombre de marcas epigenéticas. Las marcas epigenéticas permiten la persistencia de patrones estables de actividad génica, incluso en ausencia del estímulo inicial, y pueden proporcionar una «memoria» molecular de eventos medioambientales experimentados por la planta.
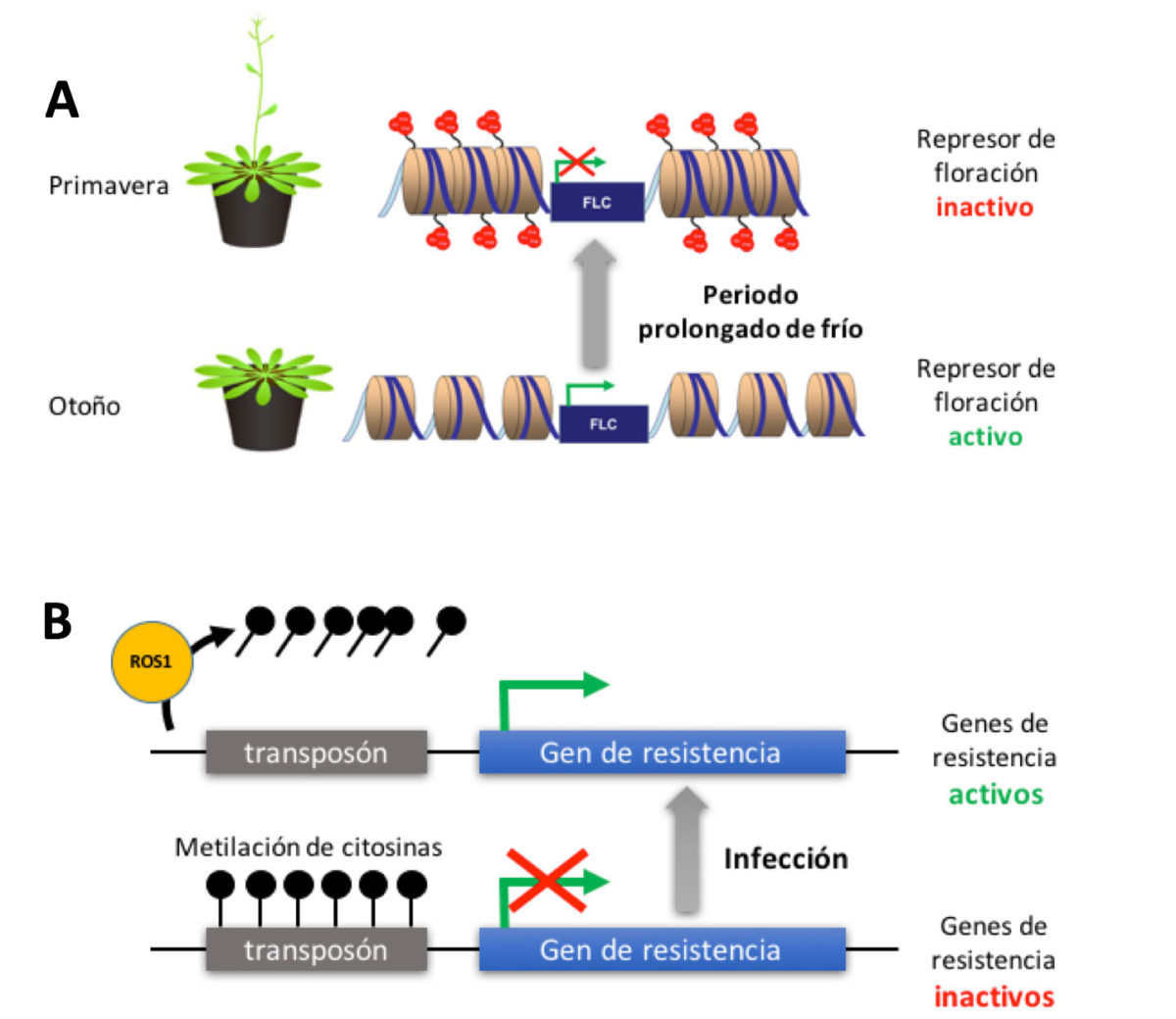
Uno de los casos mejor conocidos de control epigenético en respuesta a cambios ambientales está relacionado con el papel del frío en la floración. Para asegurar el éxito reproductor, las plantas de climas templados han de hacer coincidir su fase de floración con periodos en los que se den condiciones ambientales favorables. La clave de este ajuste reside en un proceso denominado vernalización, que impide que la planta florezca a menos que «recuerde» la experiencia de un periodo prolongado de frío. Su base molecular es el control epigenético de un represor de la floración denominado FLOWERING LOCUS C (FLC). Durante el invierno tiene lugar un silenciamiento epigenético progresivo y estable del gen FLC mediante una histona-metiltransferasa que transfiere tres grupos metilo a la lisina 27 de la histona H3. Se crea así un punto de anclaje para factores que condensan la cromatina alrededor del gen FLC, lo que «silencia» de forma estable este represor floral y permite la floración cuando llegue la primavera. Tras la floración y la formación de los frutos, en los embriones contenidos en las semillas el gen FLC vuelve a activarse, asegurando así que las plantas de la descendencia puedan experimentar a su debido tiempo el proceso de vernalización. En otras palabras, este sistema de memoria molecular del invierno se «resetea» en cada generación.
En plantas hay muchos ejemplos adicionales de alteraciones en la cromatina inducidas por cambios en el ambiente. La mayoría están relacionados con respuestas a estrés, ya sea biótico (herbívoros, agentes patógenos) o abiótico (sequía, estrés osmótico, frío intenso). En algunos casos dichas alteraciones afectan al otro componente de la cromatina: el propio ADN. La única marca epigenética detectada en el ADN hasta la fecha es la metilación de citosinas, que tiene un papel represor sobre la actividad de los genes. La metilación es llevada a cabo por ADN metiltransferasas, y en plantas las citosinas metiladas suelen encontrarse en regiones con secuencias repetitivas. Una de sus funciones es inhibir la actividad de elementos transponibles para evitar su diseminación descontrolada por el genoma, pero también participa en la regulación de muchos genes. En plantas la metilación de ADN es «borrada» por desmetilasas que extirpan las citosinas metiladas para que sean sustituidas por citosinas sin metilar, un mecanismo que no se ha detectado en animales. Estudios recientes han revelado que los procesos antagónicos de metilación y desmetilación de ADN regulan genes necesarios para la respuesta defensiva de las plantas a patógenos no virales. Estos genes están normalmente metilados y por tanto inactivos, pero tras la infección son activados por acción de la ADN-desmetilasa REPRESSOR OF SILENCING 1 (ROS1). Curiosamente, muchos de ellos contienen en su zona reguladora secuencias derivadas de transposones. Ello sugiere que en plantas algunos elementos transponibles han evolucionado para convertirse en módulos reguladores que permiten el control epigenético de genes en respuesta a estrés.
La mayoría de las modificaciones epigenéticas inducidas por el medioambiente son transitorias o se borran durante la gametogénesis y/o la embriogénesis. Sin embargo, algunas podrían persistir en los gametos y transmitirse a futuras generaciones, un proceso denominado herencia epigenética transgeneracional. Este tipo de herencia se ha propuesto también en animales, pero en plantas podría ser más frecuente debido a que desarrollan sus gametos a partir de células somáticas en la fase adulta. En teoría, esto podría facilitar que modificaciones epigenéticas adquiridas durante el crecimiento vegetativo puedan transmitirse a la descendencia. De hecho, hay estudios que sugieren la posible transmisión de una memoria de estrés en plantas. Por ejemplo, la progenie de plantas tratadas con cepas patogénicas bacterianas es más resistente a futuras infecciones que la de plantas no tratadas. Sin embargo, en este caso no ha podido demostrarse aún la implicación directa de modificaciones de la cromatina y no puede descartarse que la resistencia provenga de la acumulación de metabolitos y/o proteínas defensivas en las semillas. Por otra parte, se ha puesto en duda la posible ventaja adaptativa de transmitir a la progenie genes de defensa pre-activados, algo que supondría un coste energético considerable si el estrés no llega a repetirse. En definitiva, y al igual que en animales, por ahora no hay pruebas concluyentes de que los cambios epigenéticos inducidos por el ambiente puedan transmitirse a futuras generaciones.
REFERENCIAS
- Baulcombe, D.C., and Dean, C. (2014). Epigenetic regulation in plant responses to the environment. Cold Spring Harb Perspect Biol 6, a019471.
- Blázquez, M.A. (2011). Bases moleculares de la floración. Investigación y Ciencia, Mayo, 29-36.
- Deleris, A., Halter, T., and Navarro, L. (2016). DNA Methylation and Demethylation in Plant Immunity. Annu Rev Phytopathol 54, 579-603.
- Roldán-Arjona, T. (2010). Epigenética: entre la estabilidad del genotipo y la plasticidad del fenotipo. (https://web2020.sebbm.es/web/es/divulgacion/acercate-nuestros-cientificos/215-teresa-roldan-arjona-septiembre-2010-modificaciones-epigeneticas).
- Roldan-Arjona, T., and Ariza, R.R. (2009). DNA demethylation. Madame Curie Bioscience Database. (https://www.ncbi.nlm.nih.gov/books/NBK6365/).