Los sistemas modelo son esenciales para responder a numerosas preguntas científicas. En el caso de la Biología, disponer de organismos sencillos, con menor nivel de complejidad y organización, permite abordar el estudio en el laboratorio de numerosos procesos celulares y moleculares básicos.
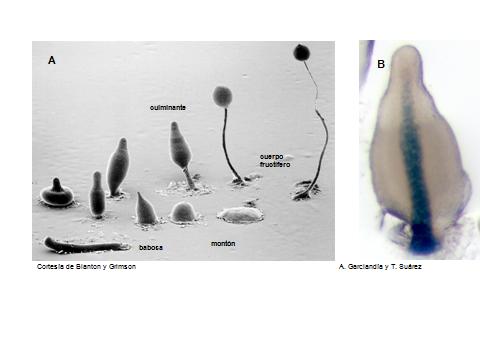
Dictyostelium discoideum, junto con Drosophila melanogaster (la mosca) o Caenorhabditis elegans (el gusano), pertenece a este selecto grupo de sistemas modelo y es, probablemente, el menos conocido de todos. Dictyostelium es una ameba, un organismo eucariota unicelular, que vive en el suelo, se alimenta de bacterias y se reproduce, sencillamente, por bipartición. Sin embargo, lo verdaderamente interesante de su ciclo de vida, es su comportamiento como ameba social. Cuando las amebas carecen de comida y las células no son capaces de dividirse, Dictyostelium opta por un ciclo de vida alternativo y se une con sus congéneres para dar lugar a una etapa de desarrollo y diferenciación. Las amebas se comunican entre sí, gracias al AMP cíclico, y 100.000 amebas se agregan para formar un montón de células. En esta agrupación, las células originalmente iguales, ya no lo son, han empezado a expresar genes diferentes unas de otras y comienza el proceso de diferenciación. Después de 24 horas, este montón de células ha dado lugar a una estructura parecida a un chupachús, tras atravesar diferentes fases: montón, babosa, culminante, cuerpo fructífero (Figura 1) (1,2). El soporte o tallo de esta estructura está formado por células muertas y la esfera, situada en la parte superior, contiene esporas capaces de resistir condiciones ambientales adversas y que originarán nuevas amebas. Las células del tallo, que son de manera sistemática un 20% del total, sufren un proceso de muerte celular programada. Es decir, este organismo, en la frontera entre lo unicelular y lo pluricelular, puede atravesar durante su ciclo de vida por etapas de diferenciación, desarrollo, morfogénesis, muerte celular,..(1,2).
En el año 2005 se completó además la secuenciación de su genoma. Esto permitió calcular que tiene 12.500 genes, casi tantos como la mosca y más de la mitad que la levadura, un organismo unicelular. El elevado número de genes es el testimonio del coste de la multicelularidad, que implica la existencia de comunicación entre células, de señales enviadas y mensajes recibidos, de mecanismos de control coordinados, todo ello imprescindible para completar una fase multicelular. Además, se ha podido comprobar que un elevado porcentaje, más de la mitad, de dichos genes están perfectamente conservados en otros organismos, incluso en mamíferos (3).
Las amebas de Dictyostelium son capaces de moverse y emplean para ello un mecanismo molecular idéntico al de los linfocitos. La distribución específica de las proteínas dentro de las células en movimiento, con las quinasas de lípidos (como PI3K) en la parte anterior de las células y las fosfatasas de lípidos (como PTEN) en la parte posterior, es similar en las amebas y en los linfocitos (4,5). Las amebas fagocitan bacterias eficazmente, empleando los mismos mecanismos que las células encargadas de esta función en organismos multicelulares.
Dictyostelium se ha empleado asimismo para estudiar los efectos secundarios de fármacos empleados en el tratamiento de cáncer. Así por ejemplo, se demostró por primera vez en este organismo que el cisplatino desequilibra los niveles de las ceramidas dentro de las células. Otro importante hito que se consiguió en Dictyostelium fue encontrar el mecanismo molecular por el que actúan los fármacos empleados como estabilizadores del ánimo (el litio o el ácido valproico) en enfermedades como el trastorno bipolar o la depresión. Se demostró, y se comprobó posteriormente en pacientes, que estos compuestos modulan las reservas de fosfolípidos como el fosfoinositol-3-fosfato (PI3P) (6).
La diferenciación celular aparece en las células en los primeros estadios del desarrollo de Dictyostelium (montón, Figura 1A); posteriormente, las células se organizan, siguiendo un patrón de desarrollo y se separan los distintos tipos celulares (Figura 1B). Durante el desarrollo de este sencillo organismo muchos de los sistemas encargados de la transducción de señales, los factores de transcripción y los receptores, están conservados en mamíferos (7). Esto permite el análisis de mutantes de desarrollo que no se pueden analizar en organismos más complejos, donde los mecanismos de compensación impiden la observación de fenotipos individuales.
La facilidad de manejo de Dictyostelium, su sencilla genética y la rapidez de crecimiento, junto con la gran cantidad de técnicas bioquímicas y de biología molecular disponibles, hacen de este organismo el sistema modelo de elección para el estudio exhaustivo de los mecanismos básicos de diferenciación celular y desarrollo.
REFERENCIAS
- Annesley SJ, Fisher PR. (2009) Dictyostelium discoideum–a model for many reasons. Mol Cell Biochem. 329:73-91.
- Urushihara H. (2009) The cellular slime mold: eukaryotic model microorganism. Exp Anim., 58:97-104.
3.Williams JG, Noegel AA, Eichinger L. (2005) Manifestations of multicellularity: Dictyostelium reports in. Trends Genet., 21:392-8. - Kim L, Kimmel AR. (2006) GSK3 at the edge: regulation of developmental specification and cell polarization. Curr Drug Targets., 7:1411-9.
- King JS, Insall RH. (2009) Chemotaxis: finding the way forward with Dictyostelium. Trends Cell Biol., 19:523-30.
- Williams RS, Boeckeler K, Gräf R, Müller-Taubenberger A, Li Z, Isberg RR, Wessels D, Soll DR, Alexander H, Alexander S. (2006) Towards a molecular understanding of human diseases using Dictyostelium discoideum. Trends Mol Med., 12:415-24.
- Williams JG. (2006) Transcriptional regulation of Dictyostelium pattern formation. EMBO Rep., 7:694-8.