
Especial Premio Fronteras del Conocimiento BBVA.
La concesión del Premio Fronteras del Conocimiento 2012 de la Fundación BBVA a Douglas Coleman y Jeffrey Friedman por el descubrimiento del gen de la leptina, hormona involucrada en la regulación central de la ingesta, supone un reconocimiento a ambos investigadores por este hallazgo crucial para la comprensión de los mecanismos reguladores del balance energético en el organismo, así como de la fisiopatología de la obesidad.
El incremento de la prevalencia de la obesidad en los países desarrollados ha incentivado la investigación sobre el papel del tejido adiposo en el desarrollo de trastornos adicionales inducidos por el aumento de peso. El tejido adiposo es un órgano endocrino que produce numerosas hormonas, llamadas adipoquinas, entre las que destaca la leptina. El descubrimiento en 1994 del gen que codifica esta hormona (1) abrió una nueva línea de conocimiento sobre el papel de los factores sintetizados por el adipocito en la regulación de la homeostasis energética y el metabolismo.
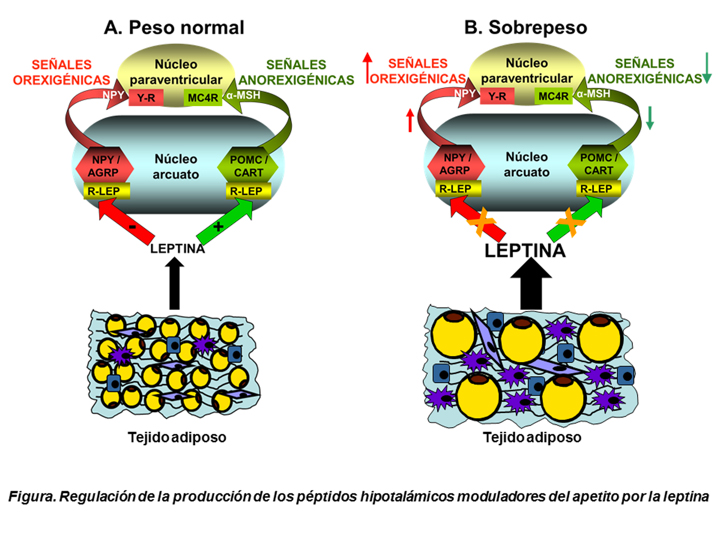
La ingesta y el control del peso corporal dependen de la interacción entre el sistema nervioso central y las señales procedentes de la periferia. La leptina, sintetizada preferentemente por el tejido adiposo, proporciona información sobre la cantidad de este tejido al hipotálamo, centro modulador de la ingesta y del balance energético (2), donde se une a sus receptores principalmente en el núcleo arcuato, una de las áreas hipotalámicas más importantes que regula esta función integradora, para inhibir la síntesis de los péptidos estimuladores del apetito denominados proteína relacionada con Agouti (AGRP) y el neuropéptido Y (NPY) y estimular la de propiomelanocortina (POMC) y el tránscrito relacionado con anfetaminas y cocaína (CART), que inhiben la ingesta (Figura, panel A). Por lo tanto, esta hormona tiene un efecto inhibidor sobre la ingesta.
La activación del receptor de leptina (R-LEP) estimula las vías de señalización intracelulares que median sus acciones, siendo muchas de las dianas intracelulares compartidas con otras hormonas, como la insulina (3). Esta hormona tiene un efecto anorexigénico similar al de la leptina, inhibiendo la síntesis de péptidos orexigénicos y estimulando la de los inhibidores del apetito. Asimismo, la leptina mejora la homeostasis de la glucosa incrementando la sensibilidad insulínica en el hipotálamo. Por lo tanto, el cruzamiento de la señalización desempeña una función relevante en la fisiología y además, los cambios en la activación de ambas vías, no solamente en el sistema nervioso, sino en la periferia, están involucrados en el desarrollo de un perfil inflamatorio desfavorable presente en diferentes enfermedades (4). Uno de los casos más frecuente de alteraciones en la señalización es la resistencia a la acción de ambas hormonas, frecuentemente por un mecanismo de inhibición por el producto final, el supresor de señalización de citoquinas 3 (SOCS3) en el caso de la leptina y la fosfotirosina fosfatasa 1B (PTP1B) en la insulina, presentando un mecanismo de inhibición cruzada. La resistencia central provoca un aumento de la síntesis de péptidos estimuladores del apetito y la inhibición de los anorexigénicos, con un balance energético positivo que se traduce en una mayor ingesta y el aumento del peso corporal (Figura, panel B).
Aunque la leptina circulante procede principalmente del adipocito, numerosos órganos periféricos y el sistema nervioso central pueden producir pequeñas cantidades, teniendo además receptores para esta adipoquina. Así, desde su descubrimiento, se ha descrito su implicación en diferentes procesos y órganos, en muchos casos relacionados con su carácter de adipoquina y sus efectos sobre la homeostasis energética. En el sistema nervioso central, no solamente el hipotálamo regula la homeostasis energética, sino que otras áreas cerebrales están involucradas en esta función reguladora. Así, la corteza cerebral y el hipocampo presentan receptores para esta hormona, que interacciona con diferentes neurotransmisores para modular funciones estrechamente conectadas con la regulación de la ingesta (5). También se han descrito acciones neuroprotectoras de la leptina en diferentes áreas cerebrales mediadas no solamente por la activación de JAK2 y STAT3, sino por dianas de la vía clásica de la insulina, como Akt, lo que viene a confirmar nuevamente la estrecha relación entre ambas hormonas.
El carácter versátil de la leptina se manifiesta por la multiplicidad de sus acciones en tejidos periféricos (6). En el sistema circulatorio, la leptina aumenta la actividad del sistema nervioso simpático, con implicaciones en la función endotelial y en la homeostasis de la presión sanguínea. Con respecto a la función respiratoria se han referido sus efectos sobre el desarrollo pulmonar, sugiriéndose un papel en el control respiratorio, Así, en obesos hay resistencia a la leptina relacionada con la apnea obstructiva. También está involucrada en la regulación del crecimiento lineal, función ligada con la homeostasis energética. De hecho, la leptina incrementa el contenido mineral óseo y tiene efectos mitogénicos sobre los condrocitos de la placa de crecimiento epifisaria, estimulando al mismo tiempo la síntesis y secreción de la hormona de crecimiento en el hipotálamo. Entre otras de sus múltiples acciones, podemos enumerar sus efectos sobre el tejido adiposo (7), aparato reproductor, riñón y su papel en la regulación de la respuesta inmune.
En resumen, la investigación sobre las vías de señalización activadas por la leptina y su interacción con otras hormonas reguladoras del metabolismo y la homeostasis energética representa un reto fascinante que ayudará a desvelar nuevos conocimientos sobre la fisiología de esta hormona y a entender los mecanismos fisiopatológicos involucrados.
Referencias:
- Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372:425-432.
- Morton GJ. Hypothalamic leptin regulation of energy homeostasis and glucose metabolism. J Physiol 2007, 583:437-443.
- Burgos-Ramos E, Chowen JA, Arilla-Ferreiro E, Canelles S, Argente J, Barrios V. Chronic central leptin infusion modifies the response to acute central insulin injection by reducing the interaction of the insulin receptor with IRS2 and increasing its association with SOCS3. J Neurochem 2011, 117:175-185.
- Martos-Moreno GÁ, Kratzsch J, Körner A, Barrios V, Hawkins F, Kiess W, Argente J. Serum visfatin and vaspin levels in prepubertal children: effect of obesity and weight loss after behavior modifications on their secretion and relationship with glucose metabolism. Int J Obes (Lond) 2011, 35:1355-1362.
- Perianes-Cachero A, Burgos-Ramos E, Puebla-Jiménez L, Canelles S, Frago LM, Hervás-Aguilar A, de Frutos S, Toledo-Lobo MV, Mela V, Viveros MP, Argente J, Chowen JA, Arilla-Ferreiro E, Barrios V. Acute up-regulation of the rat brain somatostatin receptor-effector system by leptin is related to activation of insulin signaling and may counteract central leptin actions. Neuroscience 2013, 252:289-301.
- Mantzoros CS, Magkos F, Brinkoetter M, Sienkiewicz E, Dardeno TA, Kim SY, Hamnvik OP, Koniaris A. Leptin in human physiology and pathophysiology. Am J Physiol Endocrinol Metab 2011, 301:E567-E584.
- Burgos-Ramos E, Canelles S, Perianes-Cachero A, Arilla-Ferreiro E, Argente J, Barrios V. Adipose tissue promotes a serum cytokine profile related to lower insulin sensitivity after chronic central leptin infusion. PLoS One 2012, 7:e46893.